Research
Background
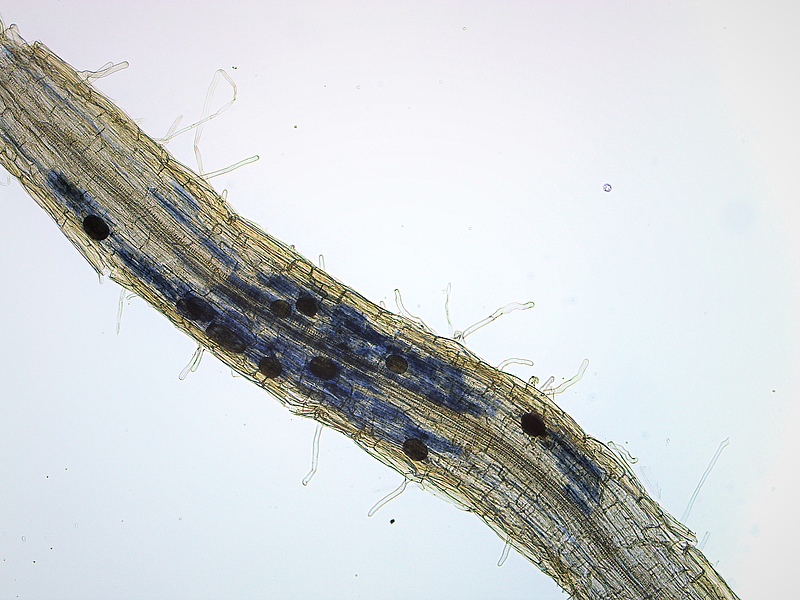
Plant mycorrhization occurs in roots in most of the terrestrial plant species and is commonly known to enhance plant growth (Smith & Read, 2010; Ferlian et al., 2018). Mycorrhizal fungi supply the plant with soil water and nutrients in exchange for photosynthetic products from the plant. As a consequence, mycorrhizal fungi have been proposed to play a critical role in positive biodiversity-ecosystem functioning relationships (Klironomos et al., 2000; Schnitzer et al., 2011). Different mycorrhizal types coexist and considerably differ in their morphological and physiological characteristics that facilitate dissimilar soil nutrient uptake processes. Associations with different mycorrhizal types may, therefore, increase complementary use of resources among plant species in diverse plant communities.
In the MyDiv Experiment, we integrate interactions between plants and mycorrhizal types into the common setting of biodiversity-ecosystem functioning experiments. We test the hypothesis that tree communities with diverse mycorrhizal types will utilise soil nutrients more complementarily than tree communities with a single mycorrhizal type. Therefore, treatments combining high tree species richness and presence of both mycorrhizal types are expected to increase resource uptake and, consequently, complementarity resulting in the highest tree performance. We manipulate the two
main mycorrhizal types (via respective tree species selection) along a tree species richness gradient.
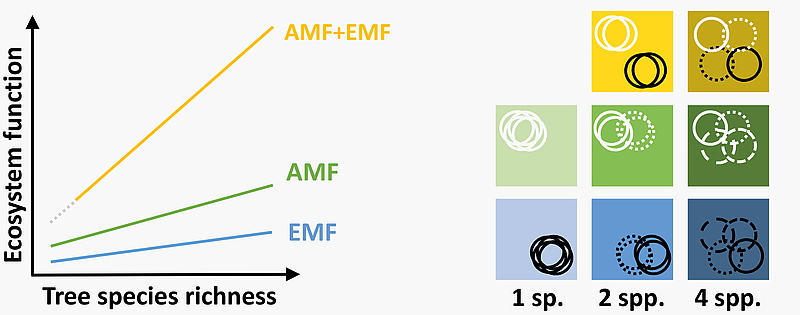
Left: hypothetical relationship between tree species richness and ecosystem functions in the different tree
communities in the MyDiv Experiment. Right: the underlying resource use scenarios in tree species that coexist in a community
and are limited by a set of resources. The boxes represent resource space; the circles represent tree species
that are different when lines are different (modified after Ferlian et al., 2018).
Long-term Measurements
After setting up the experiment, we conducted a baseline soil sampling campaign (biological, chemical, and physical soil variables) and tree measurements after the first growing season (Ferlian et al., 2018). Subsequently, most soil and tree measurements have been and will be repeated annually to create time-series data that will allow us to assess whether temporal dynamics of ecosystem functions are affected by the experimental treatments (Eisenhauer et al., 2012; Reich et al., 2012). Additionally, in the first growing season, soil cores of 1 m depth were taken from all plots and subdivided into eight layers (0-5, 5-10, 10-20, 20-30, 30-40, 40-50, 50-60, and 60-100 cm depth) and archived for future purposes, such as soil carbon dynamics and storage. The MyDiv Experiment is equipped with three climate stations spread across the site with each of them measuring air temperature and humidity as well as soil temperature and moisture at three depths in intervals of 30 minutes.

Regularly measured parameters:

Projects & Collaborations
The MyDiv Experiment is run as an open experimental platform and invites all interested researchers from iDiv’s research consortium and outside of iDiv to collaborate (see Collaboration Guidelines). Since the setup of the experiment, we have collaborated with researchers spanning a wide range of expertise in ecology, plant physiology, molecular biology, and genetics. The MyDiv Experiment also supports young scientists providing a platform for internships as well as Bachelor’s and Master’s theses (see Jobs & Theses).
Past and ongoing collaborations:

The MyDiv Experiment is further a member of the global collaborative network of tree diversity experiments called TreeDivNet. Experimental designs within this network share important characteristics, such as the manipulation of tree diversity in the field, separation of tree diversity from identity effects, a diversity gradient of at least three levels, and the assessment of multiple ecosystem functions (Ferlian et al., 2018). The network currently consists of nearly thirty experiments comprised of >1,100,000 trees across four biomes (Paquette et al. 2018). Within TreeDivNet, several global synthesis studies have evolved where the MyDiv Experiment contributed to, such as a global litter decomposition study using the Tea Bag Index (Ika Djukic, Umweltbundesamt Austria) and a global study on the effects of tree diversity on leaf herbivory in birch (Bastien Castagneyrol, Hervé Jactel, INRA, Bordeaux).


References
Eisenhauer N, Reich PB, Scheu S. 2012. Increasing plant diversity effects on productivity with time due to delayed soil biota effects on plants. Basic and Applied Ecology 13: 571-578.
Ferlian O, Cesarz S, Craven D, Hines J, Barry KE, Bruelheide H, Buscot F, Haider S, Heklau H, Herrmann S, Kühn P, Pruschitzki U, Schädler M, Wagg C, Weigelt A, Wubet T, Eisenhauer N. 2018. Mycorrhiza in tree diversity–ecosystem function relationships: conceptual framework and experimental implementation. Ecosphere 9:e02226.
Klironomos JN, McCune J, Hart M, Neville J. 2000. The influence of arbuscular mycorrhizae on the relationship between plant diversity and productivity. Ecology Letters 3: 137-141.
Paquette A, Hector A, Castagneyrol B, Vanhellemont M, Koricheva J, Scherer-Lorenzen M, Verheyen K et al. 2018. A million and more trees for science. Nature ecology & evolution 2: 763.
Reich PB, Tilman D, Isbell F, Mueller K, Hobbie SE, Flynn DF, Eisenhauer N. 2012. Impacts of biodiversity loss escalate through time as redundancy fades. Science 336: 589-592.
Schnitzer SA, Klironomos JN, HilleRisLambers J, Kinkel LL, Reich PB, Xiao K, Rillig MC, Sikes BA, Callaway RM, Mangan SA et al. 2011. Soil microbes drive the classic plant diversity-productivity pattern. Ecology 92: 296-303.
Smith SE, Read DJ. 2010. Mycorrhizal symbiosis. Academic press, London.
